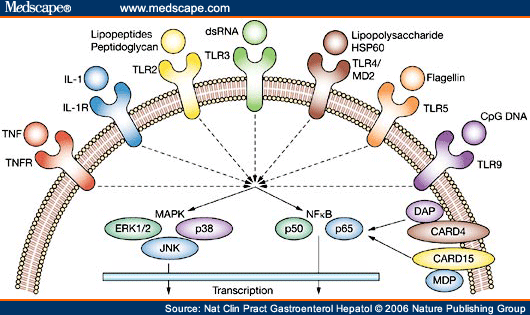
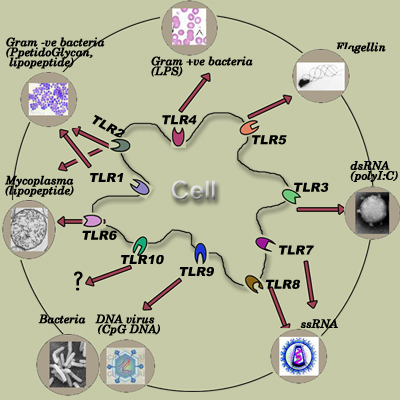
Pattern recognition receptors (PRRs) are a primitive part of the immune system. They are proteins expressed by cells of the innate immune system to identify pathogen-associated molecular patterns (PAMPs), which are associated with microbial pathogens or cellular stress, as well as damage-associated molecular patterns (DAMPs), which are associated with cell components released during cell damage. They are also called pathogen recognition receptors or primitive pattern recognition receptors because they evolved before other parts of the immune system, particularly before adaptive immunity.
Contents
- 1 Molecules recognized
- 2 Classification
- 3 PRR types and localization
- 3.1 Membrane-bound PRRs
- 3.1.1 Receptor kinases
- 3.1.2 Toll-like receptors (TLR)
- 3.1.3 C-type lectin Receptors (CLR)
- 3.1.3.1 Group I CLRs: The mannose receptors
- 3.1.3.2 Group II CLRs: asialoglycoprotein receptor family
- 3.2 Cytoplasmic PRRs
- 3.2.1 NOD-like receptors (NLR)
- 3.2.2 RIG-I-like receptors (RLR)
- 3.2.3 Plant PRRs
- 3.3 Secreted PRRs
- 4 Signaling via PRR
- 5 References
- 6 External links
Molecules recognized
The microbe-specific molecules that are recognized by a given PRR are called pathogen-associated molecular patterns (PAMPs) and include bacterial carbohydrates (such as lipopolysaccharide or LPS, mannose), nucleic acids (such as bacterial or viral DNA or RNA), bacterial peptides (flagellin, microtubule elongation factors), peptidoglycans and lipoteichoic acids (from Gram-positive bacteria), N-formylmethionine, lipoproteins and fungal glucans and chitin.
Endogenous stress signals are called danger-associated molecular patterns (DAMPs) and include uric acid.
Classification
PRRs are classified according to their ligand specificity, function, localization and/or evolutionary relationships. On the basis of function, PRRs may be divided into endocytic PRRs or signaling PRRs.
- Signaling PRRs include the large families of membrane-bound Toll-like receptors and cytoplasmic NOD-like receptors.
- Endocytic PRRs promote the attachment, engulfment and destruction of microorganisms by phagocytes, without relaying an intracellular signal. These PRRs recognize carbohydrates and include mannose receptors of macrophages, glucan receptors present on all phagocytes and scavenger receptors that recognize charged ligands, are found on all phagocytes and mediate removal of apoptotic cells.
PRR types and localization
Membrane-bound PRRs
Receptor kinases
PRRs were first discovered in plants.[1] Since that time many plant PRRs have been predicted by genomic analysis (370 in rice; 47 in Arabidopsis). Unlike animal PRRs, which associated with intracellular kinases via adaptor proteins (see non-RD kinases below), plant PRRs are composed of an extracellular domain, transmembrane domain, juxtamembrane domain and intracellular kinase domain as part of a single protein.
Toll-like receptors (TLR)
Recognition of extracellular or endosomal pathogen-associated molecular patterns is mediated by transmembrane proteins known as toll-like receptors (TLRs).[2] Toll-like receptors were first discovered in Drosophila and trigger the synthesis and secretion of cytokines and activation of other host defense programs that are necessary for innate or adaptive immune responses. TLRs have been found in many species. In mammals, these receptors have been assigned numbers 1 to 11 (TLR1-TLR11). Interaction of TLRs with their specific PAMP induces NF-κB signaling and the MAP kinase pathway and therefore the secretion of pro-inflammatory cytokines and co-stimulatory molecules. Molecules released following TLR activation signal to other cells of the immune system making TLRs key elements of innate immunity and adaptive immunity.[3]
C-type lectin Receptors (CLR)
Many different cells involved in innate immune system starting from cells as complex as immature dendritic cells to cells as small as platelets express myriad CLRs which shape innate immunity by virtue of their pattern recognition ability.[4] Most classes of human pathogens are covered by CLRs: e.g. among external or infectious pathogens mannose is the recognition motif for many viruses, fungi and mycobacteria; similarly fucose for certain bacteria and helminths; and glucans are present on mycobacteria and fungi. In addition, many of acquired nonself surfaces e.g. carcinoembryonic/oncofetal type neoantigens carrying "internal danger source"/"self turned nonself" type pathogen pattern are also identified and destroyed (e.g. by complement fixation or other cytotoxic attacks) or sequestered (phagocytosed or ensheathed) by the immune system by virtue of the CLRs. The name lectin is also a bit misleading because the family includes proteins with at least one C-type lectin domain (CTLD) which is a specific type of carbohydrate recognition domain. CTLD is a ligand binding motif found in more than 1000 known proteins (more than 100 in humans) and the ligands are often not sugars.[5] If and when the ligand is sugar they need Ca2+ - hence the name "C-type", but many of them don't even have a known sugar ligand thus despite carrying a lectin type fold structure, some of them are technically not "lectin" in function.
CLRs are often endocytic receptors but may be also signaling as well, and depending on the ligand their signaling and trafficking fate might be different. They may have direct signaling including MAP kinase and NFkB activation or may act indirectly by modulating TLRs. CLRs are a very big category and includes soluble PRRs as well (see at the end of this article) that serve many important immune functions.
Membrane receptor CLRs are broadly grouped into:
Group I CLRs: The mannose receptors
The mannose receptor (MR) is a PRR primarily present on the surface of macrophages and dendritic cells. The MR belongs to the multilectin receptor protein group and, like the TLRs, provides a link between innate and adaptive immunity.[6] It recognizes and binds to repeated mannose units on the surfaces of infectious agents and its activation triggers endocytosis and phagocytosis of the microbe via the complement system. Specifically, mannose binding triggers recruitment of MBL-associated serine proteases (MASPs). The serine proteases activate themselves in a cascade, amplifying the immune response: MBL interacts with C4, binding the C4b subunit and releasing C4a into the bloodstream; similarly, binding of C2 causes release of C2b. Together, MBL, C4b and C2a are known as the C3 convertase. C3 is cleaved into its a and b subunits, and C3b binds the convertase. These together are called the C5 convertase. Similarly again, C5b is bound and C5a is released. C5b recruits C6, C7, C8 and multiple C9s. C5, C6, C7, C8 and C9 form the membrane attack complex (MAC).
Group II CLRs: asialoglycoprotein receptor family
This is another large superfamily of CLRs that includes molecules like
- The classic asialoglycoprotein receptor macrophage galactose-type lectin (MGL)
- DC-SIGN (CLEC4L)
- Langerin (CLEC4K)
- Myeloid DAP12‑associating lectin (MDL)‑1 (CLEC5A)
- DC‑associated C‑type lectin 1 (Dectin1) subfamily which includes
- dectin 1/CLEC7A
- DNGR1/CLEC9A
- Myeloid C‑type lectin‑like receptor (MICL) (CLEC12A)
- CLEC2 (also called CLEC1B)- the platelet activation receptor for podoplanin on lymphatic endothelial cells and invading front of some carcinomas.
- CLEC12B
- DC immunoreceptor (DCIR) subfamily which includes:
- DCIR/CLEC4A
- Dectin 2/CLEC6A
- Blood DC antigen 2 (BDCA2) ( CLEC4C)
- Mincle i.e. macrophage‑inducible C‑type lectin (CLEC4E)
The nomenclature (mannose versus asialoglycoprotein) is a bit misleading as these the asialoglycoprotein receptors are not necessarily galactose (one of the commonest outer residues of asialo-glycoprotein) specific receptors and even many of this family members can also bind to mannose after which the other group is named !
Cytoplasmic PRRs
NOD-like receptors (NLR)
For more details, see NOD-like receptor.
The NOD-like receptors (NLRs) are cytoplasmic proteins that regulate inflammatory and apoptotic responses. Approximately 20 of these proteins have been found in the mammalian genome and include two major subfamilies called NODs and NALPs, the MHC Class II transactivator (CIITA), IPAF, BIRC1, and other molecules.[7] This family of proteins is greatly expanded in plants, and constitutes a core component of plant immune systems.[8] Some of these proteins recognize endogenous or microbial molecules or stress responses and form oligomers that, in animals, activate inflammatory caspases (e.g. caspase 1) causing cleavage and activation of important inflammatory cytokines such as IL-1, and/or activate the NF-κB signaling pathway to induce production of inflammatory molecules. The NLR family is known under several different names, including the CATERPILLER (or CLR) or NOD-LRR family.[7][9]
-
- NODs
- The ligands are currently known for NOD1 and NOD2. NOD1 recognizes a molecule called meso-DAP, which is a peptidoglycan constituent only of Gram negative bacteria. NOD2 proteins recognize intracellular MDP (muramyl dipeptide), which is a peptidoglycan constituent of both Gram positive and Gram negative bacteria. NODs transduce signals in the pathway of NF-κB and MAP kinases via the serine-threonine kinase called RIP2. NOD proteins are so named because they contain a nucleotide-binding oligomerization domain which binds nucleoside triphosphate. NODs signal via N-terminal CARD domains to activate downstream gene induction events, and interact with microbial molecules by means of a C-terminal leucine-rich repeat (LRR) region.[10]
-
- NALPs
- Like NODs, these proteins contain C-terminal LRRs, which appear to act as a regulatory domain and may be involved in the recognition of microbial pathogens. Also like NODs, these proteins also contain a nucleotide binding site (NBS) for nucleoside triphosphates. Interaction with other proteins (e.g. the adaptor molecule ASC) is mediated via N-terminal pyrin (PYD) domain. There are 14 members of this subfamily in humans (called NALP1 to NALP14). Mutations in NALP3 are responsible for the autoinflammatory diseases familial cold autoinflammatory syndrome, Muckle-Wells syndrome and neonatal onset multisystem inflammatory disease. Activators of NALP3 include muramyl dipeptide, bacterial DNA, ATP, toxins, double stranded RNA, paramyxoviruses and uric acid crystals. Although these specific molecules have been shown to activate NALP3, it remains unclear whether this is due to direct binding or due to cellular stress induced by these agents.
-
- Other NLRs
- Other NLRs such as IPAF and NAIP5/Birc1e have also been shown to activate caspase-1 in response to Salmonella and Legionella.
RIG-I-like receptors (RLR)
Further information: RIG-I-like receptor
Intracellular recognition of viral double-stranded (ds) and single stranded RNA has been shown to be mediated by a group of RNA Helicases which in turn recruit factors via twin N-terminal CARD domains to activate antiviral gene programs. Three such helicases have been described in mammals: RIG-I and MDA5 (recognizing 5'triphosphate-RNA and dsRNA, respectively), which activate antiviral signaling, and LGP2, which appears to act as a dominant-negative inhibitor.
-
RIG-I and Mda5-mediated signalling pathway.
Plant PRRs
Plants contain a significant number of PRRs that share remarkable structural and functional similarity with drosophila TOLL and mammalian TLRs. The first PRR identified in plants or animals was the XA21 protein, conferring resistance to the Gram-negative bacterial pathogen Xanthomonas oryzae pv. oryzae.[1][11] Since that time two other plants PRRs, Arabidopsis FLS2 (flagellin) and EFR (elongation factor Tu receptor)have been isolated.[12] The corresponding PAMPs for FLS2 and EFR have been identified.[12] Upon ligand recognition, the plant PRRs transduce "PAMP-triggered immunity" (PTI).[13] Plant immune systems also encode resistance proteins that resemble NOD-like receptors (see above), that feature NBS and LRR domains and can also carry other conserved interaction domains such as the TIR cytoplasmic domain found in Toll and Interleukin Receptors.[14] The NBS-LRR proteins are required for effector triggered immunity (ETI).
Secreted PRRs
A number of PRRs do not remain associated with the cell that produces them. Complement receptors, collectins, ficolins, pentraxins such as serum amyloid and C-reactive protein, lipid transferases, peptidoglycan recognition proteins (PGRs) and the LRR, XA21D[15] are all secreted proteins. One very important collectin is mannan-binding lectin (MBL), a major PRR of the innate immune system that binds to a wide range of bacteria, viruses, fungi and protozoa. MBL predominantly recognizes certain sugar groups on the surface of microorganisms but also binds phospholipids, nucleic acids and non-glycosylated proteins.[16] Once bound to the ligands MBL and Ficolin oligomers recruit MASP1 and MASP2 and initiate the lectin pathway of complement activation which is somewhat similar to the classical complement pathway.
Signaling via PRR
NonRD kinases
PRRs commonly associate with or contain members of a monophyletic group of kinases called the interleukin-1 receptor-associated kinase (IRAK) family that include Drosophila Pelle, human IRAKs, rice XA21 and Arabidopsis FLS2. In mammals, PRRs can also associate with members of the receptor-interacting protein (RIP) kinase family, distant relatives to the IRAK family. Some IRAK and RIP family kinases fall into a small functional class of kinases termed non-RD, many of which do not autophosphorylate the activation loop. A survey of the yeast, fly, worm, human, Arabidopsis, and rice kinomes (3,723 kinases) revealed that despite the small number of non-RD kinases in these genomes (9%–29%), 12 of 15 kinases known or predicted to function in PRR signaling fall into the non-RD class.[17] In plants, all PRRs characterized to date belong to the non-RD class. These data indicate that kinases associated with PRRs can largely be predicted by the lack of a single conserved residue and reveal new potential plant PRR subfamilies.
References
- ^ a b Song WY, Wang GL, Chen LL, Kim HS, Pi LY, Holsten T, Gardner J, Wang B, Zhai WX, Zhu LH, Fauquet C, Ronald P (December 1995). "A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21". Science 270 (5243): 1804–6. Bibcode:1995Sci...270.1804S. doi:10.1126/science.270.5243.1804. PMID 8525370.
- ^ Beutler B, Jiang Z, Georgel P, Crozat K, Croker B, Rutschmann S, Du X, Hoebe K (2006). "Genetic analysis of host resistance: Toll-like receptor signaling and immunity at large". Annu. Rev. Immunol. 24: 353–389. doi:10.1146/annurev.immunol.24.021605.090552. PMID 16551253.
- ^ Doyle SL, O'Neill LA (October 2006). "Toll-like receptors: from the discovery of NFkappaB to new insights into transcriptional regulations in innate immunity". Biochem. Pharmacol. 72 (9): 1102–1113. doi:10.1016/j.bcp.2006.07.010. PMID 16930560.
- ^ Nat Rev Immunol. 2009 Jul;9(7):465-79. Signalling through C-type lectin receptors: shaping immune responses. Geijtenbeek TB, Gringhuis SI. http://www.mh-hannover.de/fileadmin/mhh/bilder/international/hbrs_mdphd/ZIB/Vorlesungen/Paper_09-10/Rev_IM_Geijtenbeek.pdf
- ^ Cummings RD, McEver RP. C-type Lectins. In: Varki A, Cummings RD, Esko JD, et al., editors. Essentials of Glycobiology. 2nd edition. Cold Spring Harbor (NY): Cold Spring Harbor Laboratory Press; 2009. http://www.ncbi.nlm.nih.gov/books/NBK1943/
- ^ Apostolopoulos V, McKenzie IF (September 2001). "Role of the mannose receptor in the immune response". Curr. Mol. Med. 1 (4): 469–74. doi:10.2174/1566524013363645. PMID 11899091.
- ^ a b Ting JP, Williams KL (April 2005). "The CATERPILLER family: an ancient family of immune/apoptotic proteins". Clin. Immunol. 115 (1): 33–37. doi:10.1016/j.clim.2005.02.007. PMID 15870018.
- ^ Jones DG, Dangl JL (2006). "The plant immune system". Nature 444 (7117): 323–329. Bibcode:2006Natur.444..323J. doi:10.1038/nature05286. PMID 17108957.
- ^ Inohara, Inohara, McDonald C, Nuñez G (2005). "NOD-LRR proteins: role in host-microbial interactions and inflammatory disease". Annu. Rev. Biochem. 74: 355–383. doi:10.1146/annurev.biochem.74.082803.133347. PMID 15952891.
- ^ Strober W, Murray PJ, Kitani A, Watanabe T (January 2006). "Signalling pathways and molecular interactions of NOD1 and NOD2". Nat. Rev. Immunol. 6 (1): 9–20. doi:10.1038/nri1747. PMID 16493424.
- ^ Bahar O, Pruitt R, Luu DD, Schwessinger B, Daudi A, Liu F, Ruan R, Fontaine-Bodin L, Koebnik R, Ronald P (2014). "The Xanthomonas Ax21 protein is processed by the general secretory system and is secreted in association with outer membrane vesicles". PeerJ (2). doi:10.7717/peerj.242. PMID 24482761.
- ^ a b Boller T, Felix G (2009). "A renaissance of elicitors: perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors". Annu Rev Plant Biol 60: 379–406. doi:10.1146/annurev.arplant.57.032905.105346. PMID 19400727.
- ^ Chisholm ST, Coaker G, Day B, Staskawicz BJ (2006). "Host-microbe interactions: shaping the evolution of the plant immune response". Cell 124 (4): 803–814. doi:10.1016/j.cell.2006.02.008. PMID 16497589.
- ^ McHale L, Tan X, Koehl P, Michelmore RW (2006). "Plant NBS-LRR proteins: adaptable guards". Genome Biol 7 (4): 212. doi:10.1186/gb-2006-7-4-212. PMC 1557992. PMID 16677430.
- ^ Wang GL, Ruan DL, Song WY, Sideris S, Chen L, Pi LY, Zhang S, Zhang Z, Fauquet C, Gaut BS, Whalen MC, Ronald PC (May 1998). "Xa21D encodes a receptor-like molecule with a leucine-rich repeat domain that determines race-specific recognition and is subject to adaptive evolution". Plant Cell 10 (5): 765–79. doi:10.2307/3870663. PMC 144027. PMID 9596635.
- ^ Dommett RM, Klein N, Turner MW (September 2006). "Mannose-binding lectin in innate immunity: past, present and future". Tissue Antigens 68 (3): 193–209. doi:10.1111/j.1399-0039.2006.00649.x. PMID 16948640.
- ^ Dardick C, Ronald P (January 2006). "Plant and animal pathogen recognition receptors signal through non-RD kinases". PLoS Pathog. 2 (1): e2. doi:10.1371/journal.ppat.0020002. PMC 1331981. PMID 16424920.
External links
- Pattern Recognition Receptors at the US National Library of Medicine Medical Subject Headings (MeSH)
|
Innate immune system: Pattern recognition receptors
|
|
| Membrane-bound_PRRs |
- Toll-like receptor
- Mannose receptor
- Formyl peptide receptor
- Scavenger receptor
|
|
| Cytoplasmic_PRRs |
- Toll-like receptor
- NOD-like receptor
- RIG-I-like receptor
- Scavenger receptor
|
|
| Secreted_PRRs |
- Complement receptor
- Collectin
|
|
| Other/ungrouped |
- Immunophilins (Cyclophilin)
|
|
|
Index of cells from bone marrow
|
|
| Description |
- Immune system
- Cells
- Physiology
- coagulation
- proteins
- granule contents
- colony-stimulating
- heme and porphyrin
|
|
| Disease |
- Red blood cell
- Monocyte and granulocyte
- Neoplasms and cancer
- Histiocytosis
- Symptoms and signs
- Blood tests
|
|
| Treatment |
- Transfusion
- Drugs
- thrombosis
- bleeding
- other
|
|
|