WordNet
- of or belonging to neither the right nor the left politically or intellectually
- a building dedicated to a particular activity; "they were raising money to build a new center for research" (同)centre
- (American football) the position of the player on the line of scrimmage who puts the ball in play; "it is a centers responsibility to get the football to the quarterback"
- a position on a basketball team of the player who participates in the jump that starts the game
- the position on a hockey team of the player who participates in the face off at the beginning of the game
- a place where some particular activity is concentrated; "they received messages from several centers" (同)centre
- the sweet central portion of a piece of candy that is enclosed in chocolate or some other covering (同)centre
- the object upon which interest and attention focuses; "his stories made him the center of the party" (同)centre, center of attention, centre of attention
- an area that is approximately central within some larger region; "it is in the center of town"; "they ran forward into the heart of the struggle"; "they were in the eye of the storm" (同)centre, middle, heart, eye
- a point equidistant from the ends of a line or the extremities of a figure (同)centre, midpoint
- a cluster of nerve cells governing a specific bodily process; "in most people the speech center is in the left hemisphere" (同)centre, nerve_center, nerve centre
- (football) the person who plays center on the line of scrimmage and snaps the ball to the quarterback; "the center fumbled the handoff" (同)snapper
- (basketball) the person who plays center on a basketball team
- (ice hockey) the person who plays center on a hockey team
- politically moderate persons; centrists
- the middle of a military or naval formation; "they had to reinforce the center"
- move into the center; "That vase in the picture is not centered" (同)centre
- equally distant from the extremes (同)halfway, middle, midway
- a fractional monetary unit of several countries
- (American football) putting the ball in play by passing it (between the legs) to a back; "the quarterback fumbled the snap" (同)snap
- being or placed in the center
- a microscopically small tubule
PrepTutorEJDIC
- 〈C〉『セント』(1ドルの1/100;米国・カナダの貨幣単位;《略》¢) / 〈C〉1セント銅貨 / 〈U〉(単位としての)100
- 中軸(中心)を持つ
Wikipedia preview
出典(authority):フリー百科事典『ウィキペディア(Wikipedia)』「2012/11/23 17:09:41」(JST)
wiki en
UpToDate Contents
全文を閲覧するには購読必要です。 To read the full text you will need to subscribe.
- 1. 特発性器質化肺炎 cryptogenic organizing pneumonia
- 2. 濾胞性リンパ腫の臨床症状、病理学的特徴、診断、および予後 clinical manifestations pathologic features diagnosis and prognosis of follicular lymphoma
- 3. 原発性皮膚濾胞中心リンパ腫 primary cutaneous follicle center lymphoma
- 4. 間質性肺疾患における肺生検結果の解釈 interpretation of lung biopsy results in interstitial lung disease
- 5. 特発性間質性肺炎:臨床症状および病理 idiopathic interstitial pneumonias clinical manifestations and pathology
English Journal
- Nezha/CAMSAP3 and CAMSAP2 cooperate in epithelial-specific organization of noncentrosomal microtubules.
- Tanaka N, Meng W, Nagae S, Takeichi M.SourceRIKEN Center for Developmental Biology, Kobe 650-0047, Japan.
- Proceedings of the National Academy of Sciences of the United States of America.Proc Natl Acad Sci U S A.2012 Nov 19. [Epub ahead of print]
- Major microtubules in epithelial cells are not anchored to the centrosome, in contrast to the centrosomal radiation of microtubules in other cell types. It remains to be discovered how these epithelial microtubules are generated and stabilized at noncentrosomal sites. Here, we found that Nezha [also
- PMID 23169647
- Microtubule dynamics alter the interphase nucleus.
- Gerlitz G, Reiner O, Bustin M.SourceProtein Section, Laboratory of Metabolism, National Cancer Institute, US National Institutes of Health, Bethesda, MD, 20892, USA, gabi.gerlitz@gmail.com.
- Cellular and molecular life sciences : CMLS.Cell Mol Life Sci.2012 Nov 2. [Epub ahead of print]
- Microtubules are known to drive chromosome movements and to induce nuclear envelope breakdown during mitosis and meiosis. Here we show that microtubules can enforce nuclear envelope folding and alter the levels of nuclear envelope-associated heterochromatin during interphase, when the nuclear envelo
- PMID 23117601
Japanese Journal
- Dynamic Change of Cellular Localization of Microtubule-Organizing Center During Conjugation of Ciliate Tetrahymena thermophila
- Kushida Yasuharu,Takaine Masak,Nakano Kentaro [他]
- Zoological science 32(1), 25-32, 2015-01
- NAID 40020363740
- Functional implications of the Golgi and microtubular network in gonadotropes.
- 渡部 剛,暮地本 宙己,Koga Daisuke,Hosaka Masahiro,牛木 辰男
- Molecular and Cellular Endocrinology 385(1-2), 88-96, 2014-03
- … At the center of the spherical Golgi, a pair of centrioles is situated as a microtubule-organizing center from which radiating microtubules isotropically extend toward the cell periphery. …
- NAID 120005496543
- YB-1 Functions as a Porter To Lead Influenza Virus Ribonucleoprotein Complexes to Microtubules
- Kawaguchi Atsushi,Matsumoto Ken,Nagata Kyosuke
- Journal of virology 86(20), 11086-11095, 2012-10
- … After nuclear export, using YB-1 knockdown cells and in vitro reconstituted systems, YB-1 was shown to be required for the interaction of vRNP exported from the nucleus with microtubules around the microtubule-organizing center (MTOC), where Rab11a-positive recycling endosomes were located. …
- NAID 120005399279
Related Links
- The European Bioinformatics Institute ... Database Code Database Name UniProtKB Universal Protein Knowledgebase UniProtKB/Swiss-Prot UniProtKB/Swiss-Prot UniProtKB/TrEMBL UniProtKB-TrEMBL protein sequence database
- Centrosomes function primarily as microtubule-organizing centres and play a crucial role during mitosis by organizing the bipolar spindle. In addition to this function, centrosomes act as reaction centers where numerous key regulators ...
Related Pictures




★リンクテーブル★
| リンク元 | 「MTOC」「微小管重合中心」 |
| 関連記事 | 「center」 |
「MTOC」
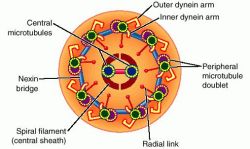
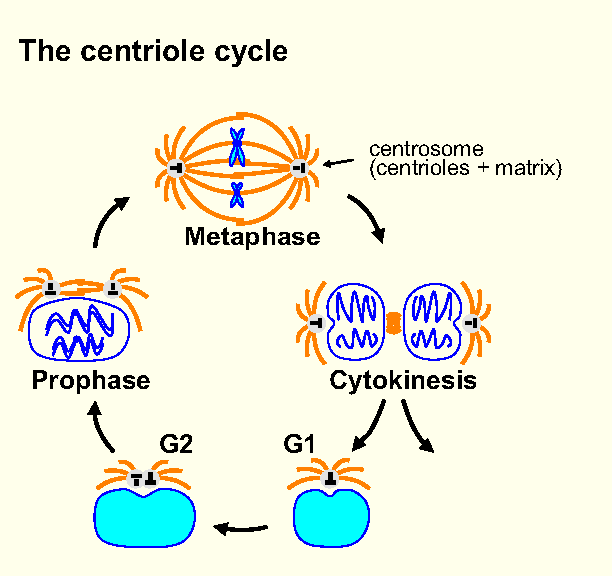
[★] 微小管重合中心 microtubule-organizing center
「微小管重合中心」
「center」
- n.