|
|
この記事は検証可能な参考文献や出典が全く示されていないか、不十分です。
出典を追加して記事の信頼性向上にご協力ください。(2013年1月) |
視覚(しかく、英: vision[1][2])とは、眼を受容器とする感覚のこと[3]。
目次
- 1 概説
- 2 視覚の研究史
- 3 視覚情報処理
- 3.1 視覚刺激
- 3.2 視感度と錐体分光感度
- 3.3 視野
- 3.4 時空間特性
- 3.4.1 空間周波数特性と視力
- 3.4.2 空間周波数チャネル
- 3.4.3 時間周波数特性とCFF
- 3.5 明るさ
- 3.6 形
- 3.7 奥行きの知覚
- 4 視覚神経科学
- 4.1 網膜における情報処理
- 4.2 皮質下における処理
- 4.3 皮質における処理
- 5 視覚における困難
- 6 進化と色覚
- 7 脚注
- 8 参考文献
- 9 関連項目
- 10 外部リンク
概説
いわゆる五感のひとつである。
視覚とは、光のエネルギーが網膜上の感覚細胞に対する刺激となって生じる感覚のことである[3]。「視覚」という言葉は、形態覚、運動覚、色覚、明暗覚などの総称として用いられている[3]。
視覚によって、外界にある物体の色、形、運動、テクスチャ、奥行きなどについての情報、物体のカテゴリーについての情報[要出典]、物体の位置関係のような外界の空間的な情報などが得られる。
「したがって、視覚は光情報をもとに外界の構造を推定する過程とみなせる」と言う[誰?]。脊椎動物の神経系では、可視光は網膜において符号化され、外側膝状体 (LGN) を経て大脳皮質において処理される。[4]
本稿ではヒトを中心に、動物の視覚のみを扱う。脊椎動物(ヒトを含む)、節足動物(昆虫、甲殻類)、軟体動物(タコ、イカ)など、多くの動物が視覚をもつ。
なお、視覚を用いて認識することを「見る(みる)」といい[5]、転じて、「読む」、「会う」、「試す」などの意味もある(「試す」の意味での「見る」は、一般的には仮名書きされる)。遠くから大局を眺める、というニュアンスや、あるいは、深い認識の過程(いわゆる「心の目」)のほうを積極的に使う、といったニュアンスを含む場合は「観る」とも書く。
視覚の研究史
前史
プラトンは視覚を聴覚とともに、対象から離れても成立するため、他の感覚より優れたものと位置付けた。西洋ではこの見解が継承され、伝統的に、視覚および聴覚に関わるもののみが芸術とみなされてきた。
イギリス経験論では、視覚は他の感覚入力との連合によって説明された。経験論哲学における有名な問題として、「球体と立方体を触覚的に判別できる先天盲者が開眼手術を受けたとき、盲人は視覚的に球体と立方体を判別できるか」というモリニュクス問題がある。経験論によれば、視覚は他の感覚と連合されていないため、開眼時点では視覚的な判断はできないと結論された。ヘルムホルツは視覚を感覚入力をもとにした無意識的推論の過程であると見なした。例えば、小さなものや遠くにあるものは、網膜上では同じように小さく見える。しかし、我々は小さな顔を見たとしても、顔が小さいと知覚することはなく、顔が遠くにあるように知覚する(大きさの恒常性)。このことは、「顔というものは実際にはこの程度の大きさのはずだから、網膜上で顔が小さいということは遠くにあるのだろう」という推論を我々が無意識的に行っているのだと解釈された。
神経科学
1866年にシュルツは固定染色法により、形態的に異なる2種類の光受容器(桿体と錐体)があることを確認した。1930年代にハートラインは単一視神経の光応答をカブトガニより初めて測定し、受容野の概念を提唱した。1950年代にクフラーは網膜神経節細胞が拮抗的受容野を持つことを発見した。1950年代後半にヒューベルとヴィーゼルは、大脳皮質の神経細胞は線分などの特徴をもつ刺激に対して選択的に応答することを報告した。また、発達期に視覚刺激の入力が遮断されると、遮断された刺激に対して選択的応答を示す神経細胞の数は減少することを報告した。1970年代後半になりパッチクランプ法が開発されると、視細胞の光受容機構の研究が進んだ。1990年代には脳機能イメージング技術が進展した。
視覚情報処理
光学系を通じて網膜に投影される網膜像は、三次元世界の物理法則である光学によって決定される。視覚は、網膜像をもとに外界の三次元構造を復元する情報処理とみなせる。そのため、光学によって三次元世界の構造から網膜像が生じるのに対して、視覚は網膜像から外界の三次元構造の推定という逆問題を解いていることになる。このことから、視覚情報処理は逆光学とよばれる。ところが、光学は三次元の外界から二次元の網膜像への対応を決定するため、網膜平面に対して奥行き方向の情報は、網膜像では完全に失われてしまう。したがって、網膜像から外界の構造復元という逆問題は、そもそも理論的に解くことのできない問題である。そのため、視覚情報処理は不良設定問題である。おおまかには不良設定問題は、正しい解を一意に求めることができない問題のことである。不良設定問題は、何らかの制約条件を設けなければ解くことができない。視覚系は外界の構造に関するさまざまな仮定を設けることで、逆問題を解いている。ところが、そもそも視覚情報処理は不良設定であるため、こうした仮定が常に正しいとは限らない。そのため、視覚系が用いている外界についての仮定が、物理的世界での規則と異なっていた場合には、物理的世界の構造を反映しない知覚が得られることになる(錯視)。
網膜像は、外界の構造、光源の位置と性質、観察者と外界の位置関係などによって変化する。ところが多くの場合では、網膜像の変化にも関わらず、外界の構造を反映する一定した知覚が得られる。視覚のこのような性質を恒常性とよぶ。たとえば照明光の光量が変化して網膜像における平均輝度が上昇しても、物体表面の明るさの知覚は変化しない(明るさの恒常性)。あるいは、物体の網膜像における大きさは、物体と観察者との距離(観察距離)に応じて変化する。しかし、知覚される物体の大きさは、観察距離の影響を受けにくい(大きさの恒常性)。このように、視覚では近刺激そのものの物理的性質が知覚されるのではなく、遠刺激の性質を反映した知覚が得られる。
視覚刺激
物体が網膜において結ぶ像の大きさを、視角によって表現する。視角とは物体の両端から結点に引いた線のなす角度のことである。中心窩からの視角を偏心度とよぶ。視覚系に入力した画像の各点の性質は、輝度と色によって記述される。輝度と色は、画像の一点のみで決定できる視覚属性であるため、一次属性とよぶ。テクスチャ、運動、両眼視差のように、空間的・時間的に異なる画像の複数の点において定義される視覚属性を、二次属性あるいは高次属性とよぶ。網膜像が空間的周期をもつとき、周期の細かさを空間周波数によって記述する。空間周波数の単位は、c/d(cycle per degree; 視角1度あたりの周期)をとることが多い。時間的周期については、Hzが用いられる。視覚刺激を記述する際には、輝度コントラストの定義として 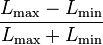 を用いることが多い。とは、画像中の輝度値の最大値と最小値を表す。この定義をMichelsonコントラストとよぶ。
を用いることが多い。とは、画像中の輝度値の最大値と最小値を表す。この定義をMichelsonコントラストとよぶ。
視感度と錐体分光感度
人間の錐体細胞(S, M, L)と桿体細胞(R)が含む視物質の吸収スペクトル
詳細は「視感度」を参照
視覚系の感度は、光の波長によって異なる。ヒト視覚系の視感度は、明所視では555 nmでピーク値をとる。このときの感度を基準として、他の波長の光に対する感度を求めると、可視光全体に対する比視感度が求まる。暗所視では507 nmの光に対して最も感度がよい。暗所では感度曲線が短波長側にシフトしている。この事実をプルキンエシフトとよぶ。放射輝度と視感度をかけ合わせた値を輝度とよぶ。
明所視では色が知覚される。色覚異常者の視感度曲線や等色関数から、分光感度の異なる3種類の光受容器(錐体)が存在することが示唆される(三色説)。健常者の等色関数および2色型色覚異常者の混同色中心から、錐体分光感度を求めることができる。暗所視における光受容器(桿体)は1種類であるため色覚は存在しない。桿体分光感度は暗所視視感度に等しい。
視野
詳細は「視野」を参照
視野とは、視覚刺激が処理できる視角の大きさである。視野は中心窩を基準として測定する。視野の大きさは動物種によって異なる。ヒト健常者の視野は、垂直方向に上側60度、下側75度程度である。水平方向では、単眼の場合、鼻側60度、耳側100度程度である。したがって、両眼で重複する視野が120度程度存在する。このことにより両眼視差が生じており、両眼立体視に寄与している。中心窩を基準に、左右や上下の領域を、左視野、上視野のように呼ぶ。各眼の耳側15度程度の位置に盲点が存在する。中心窩から20度程度の領域を中心視野とよぶ。それ以外の領域を周辺視野とよぶ。一般に中心視野ほど空間分解能が高い。周辺視野では色覚が失われる。視覚障害者(ロービジョン)には、視野欠損を示す者が含まれる。
時空間特性
空間周波数特性と視力
視覚系のコントラスト感度を空間周波数ごとに調べたものをコントラスト感度関数 (Contrast Sensitivity Function; CSF) とよぶ。静止刺激に対するヒトのCSFはバンドパス型であり、6 cpd付近で感度が最大になる。低空間周波数での感度低下は神経的原因に由来する。高空間周波数では60 cpdまで感度をもつ。高空間周波数での感度低下は主として光学的原因に由来する。一般にCSFを測定するのは煩雑であるため、光学的異常の検査目的には簡便な視力検査を行う。おおまかには視力は一定のコントラストのもとで刺激が検出できる最大の空間周波数に相当する。
空間周波数チャネル
CSFは単一の機構に由来するのではなく、複数のバンドパス型チャネルによって構成されることが分かっている。各々のチャネルはバンド幅が等しく中心周波数が異なる。チャネルは画像中の空間周波数成分の検出をしているとみなせることから、これらのチャネルを空間周波数チャネルとよぶ。空間周波数は視野ごとに存在すると考えられている。そのため、空間周波数チャネルによる処理は、大局的フーリエ変換のような線型変換ではなく、擬線型な過程とみなせる。
時間周波数特性とCFF
視覚系のコントラスト感度を時間周波数ごとに調べたものを時間的CSFとよぶ。低空間周波数では、CSFは低時間周波数で感度が低下するバンドパス型である。高空間周波数では、ローパス型である。刺激をコントラスト反転したときにフリッカーが知覚されなくなる時間周波数を臨界融合周波数 (Critical Flicker Frequency; CFF) とよぶ。CFFは一定のコントラストのもとで刺激が検出できる最大の時間周波数に相当する。ヒトのCFFは50 Hz程度とされる。
明るさ
- ヴェーバー‐フェヒナーの法則
- マッハバンド
- ヘルマン格子
形
奥行きの知覚
網膜は面であるため、網膜に投影される像は二次元である。しかし、人間は三次元空間を知覚している。これは人間が様々な奥行き手がかりをもとに、二次元情報から三次元情報への推定を行っているためである。奥行きの手がかりとして、以下のものが挙げられる。
- 単眼性のもの
- 絶対距離
- 相対距離
- 網膜像の大きさ(大きいものほど近い)
- 相対位置(上にあるものは遠く、下にあるものは近い)
- 重なり(遮蔽されているものが奥にある)
- 線遠近法
- 運動視差
- 空気遠近(遠いものほど色の差が乏しくなり、場合によっては更に青色がかる)
- 明暗関係(バルール)
- 色合い(進出色と後退色)
- 両眼性のもの(単眼性と重複するものは省略)
視覚神経科学
背側皮質視覚路が緑、腹側皮質視覚路が紫で示されている。
眼は、感覚器のひとつであり、角膜などの光学系と神経系の一部である網膜から構成される。光学系は角膜、水晶体、瞳孔などから構成され、光量の調整や焦点の調節などの機能をもつ。網膜の光受容細胞では光の強度と波長の分布が神経信号に符号化される。網膜において符号化された情報は、神経細胞の間を神経線維の興奮の伝導の形で伝えられる。以降の一連の神経線維の経路を視覚路と呼ぶ。反対色などの視覚特性は網膜内での処理に由来すると考えられている。網膜からは視神経が発しており、外側膝状体 (LGN) に投射している。外側膝状体からは視覚野への投射がある。視神経は上丘や視床の一部にも投射するが、こうした情報伝導路は眼球運動や概日周期などの非画像的な情報処理に関与するものであり、視覚情報処理の主たる経路は外側膝状体から第一次視覚野に至る経路であると考えられている。第一次視覚野からは、それ以降の高次視覚野に対して投射が存在する。第一次視覚野以降の伝導路は、物体の形状や色を処理する"What"経路と、物体の空間における位置や運動を処理する"Where"経路に二分される。"What"経路は後頭葉から側頭葉に向かい、腹側皮質視覚路と呼ばれる。"Where"経路は後頭葉から頭頂葉に向かい、背側皮質視覚路と呼ばれる。こうして処理された情報は、前頭葉などでさらに高次な処理を受けている可能性がある。
網膜における情報処理
光学系を通過した光は、網膜において網膜像を結ぶ。網膜像は網膜上の光受容細胞によってサンプリングされ、神経信号として符号化される。光受容細胞には分光感度特性の異なる桿体および錐体がある。光受容細胞は(ヒトの場合)1億個以上存在する。錐体は網膜の中心部(黄斑と呼ばれる)で密に分布し、桿体は中心部に少なく周辺部に多く分布している。光受容細胞は光入力に対して、電気的な信号によって応答する。光受容細胞の応答は、網膜内の神経節細胞に伝わる。神経節細胞の軸索は片眼で100万本程度あり、視神経乳頭より束になって眼球を出て、左右の視神経を形成し、さらに間脳の腹側から脳内に進み、間脳の視床の一部である外側膝状体(または外側膝状核)と呼ばれる神経核に達する。そこで、外側膝状体の神経細胞とシナプスを形成する。
皮質下における処理
皮質における処理
詳細は「視覚野」を参照
外側膝状体の神経細胞の軸索は大脳新皮質の後頭葉の一次視覚野に達する。
視覚における困難
- 不良設定問題
- 結合問題
- 窓問題
- ホムンクルス
- おばあさん細胞
- クオリア
進化と色覚
脊椎動物の色覚は、網膜の中にどのタイプの錐体細胞を持つかによって決まる。魚類、両生類、爬虫類、鳥類には4タイプの錐体細胞(4色型色覚)を持つものが多い。よってこれらの生物は長波長域から短波長域である近紫外線までを認識できるものと考えられている。一方ほとんどの哺乳類は錐体細胞を2タイプ(2色型色覚)しか持たない。哺乳類の祖先である爬虫類は4タイプ全ての錐体細胞を持っていたが、2億2500万年前には、最初の哺乳類と言われるアデロバシレウスが生息し始め、初期の哺乳類は主に夜行性であったため、色覚は生存に必須ではなかった。結果、4タイプのうち2タイプの錐体細胞を失い、青を中心に感知するS錐体と赤を中心に感知するL錐体の2錐体のみを保有するに至った。これは赤と緑を十分に区別できないいわゆる「赤緑色盲」の状態である。この色覚が哺乳類の子孫に遺伝的に受け継がれることとなった[6]。
霊長類真猿下目の狭鼻下目(旧世界サル)と広鼻下目(新世界サル)とが分岐したのは3000-4000万年前と言われている[7][8][要高次出典]。ヒトを含む旧世界の霊長類(狭鼻下目)の祖先は、約3000万年前、X染色体にL錐体から変異した緑を中心に感知する新たなタイプの錐体(M錐体)視物質の遺伝子が出現し、ヘテロ接合体の2本のX染色体を持つメスのみが3色型色覚を有するようになり、さらにヘテロ接合体のメスにおいて相同組換えによる遺伝子重複の変異が起こり、同一のX染色体上に2タイプの錐体視物質の遺伝子が保持されることとなり、X染色体を1本しか持たないオスも3色型色覚を有するようになった。これによって、第3の錐体細胞が「再生」された。3色型色覚は果実等の発見に有利だったと考えられる[6][7]。
時代を下ってヒトの色覚を鑑みるに、ヒトが属する狭鼻下目のマカクザルに色盲がヒトよりも非常に少ないことを考慮すると、ヒトの祖先が狩猟生活をするようになり3色型色覚の優位性が低くなり、2色型色覚の淘汰圧が下がったと考えられる[6]。色盲の出現頻度は狭鼻下目のカニクイザルで0.4%、チンパンジーで1.7%である[7]。広鼻下目のヨザルは1色型色覚であり、ホエザルは狭鼻下目と同様に3色型色覚を再獲得している[9][要高次出典]が、これらを除き、残りの新世界ザル(広鼻下目)はヘテロ接合体のX染色体を2本持つメスのみが3色型色覚を有し、オスは全て色盲である。これは狭鼻下目のようなX染色体上での相同組換えによる遺伝子重複の変異を起こさなかったためである[7]。ヒトは上記のような霊長目狭鼻下目の祖先のX染色体の遺伝子変異を受け継いでいるため、M錐体を欠損したX染色体に関連する赤緑色盲が伴性劣性遺伝をする。男性ではX染色体の赤緑色盲の遺伝子を受け継いでいると色盲が発現し、女性では2本のX染色体とも赤緑色盲の遺伝子を受け継いでいる場合に色盲が発現する[10]。
脚注
- ^ 文部省 『学術用語集 心理学編』 日本心理学会、日本学術振興会、1986年。ISBN 4-8181-8602-3。
- ^ 文部省 『学術用語集 動物学編』 日本動物学会、丸善、1988年、増訂版。ISBN 4-621-03256-9。
- ^ a b c 『広辞苑』 新村 出、岩波書店、1998年、第5版、1141頁。ISBN 978-4000801119。
- ^ コンピュータビジョンでは、光センサーからの光情報の入力をもとにした処理が行われる。
- ^ 『広辞苑』 新村 出、岩波書店、1998年、第5版、2582頁。ISBN 978-4000801119。
- ^ a b c 岡部正隆、伊藤啓「なぜ赤オプシン遺伝子と緑オプシン遺伝子が並んで配置しているのか」、『細胞工学』第21巻第7号、2002年7月。
- ^ a b c d 三上章允 (2004年9月18日). “霊長類の色覚と進化 (PDF)”. 公開講座「遺伝子から社会まで」. 京都大学霊長類研究所. 2013年9月20日閲覧。
- ^ Surridge et al. Trends Ecol. Evol. 18, 198-205, 2003
- ^ 研究の背景[リンク切れ]
- ^ 岡部正隆、伊藤啓「女性で赤緑色盲が少ない理由」、『細胞工学』第21巻第7号、2002年7月。
参考文献
|
出典は列挙するだけでなく、脚注などを用いてどの記述の情報源であるかを明示してください。記事の信頼性向上にご協力をお願いいたします。(2012年12月) |
- リチャード・L.グレゴリー 『脳と視覚 : グレゴリーの視覚心理学』 近藤倫明・中溝幸夫・三浦佳世訳、ブレーン出版、2001年。ISBN 4-89242-664-4。
- サイモン・イングス 『見る : 眼の誕生はわたしたちをどう変えたか』 吉田利子訳、早川書房、2009年。ISBN 978-4-15-208999-1。
関連項目
|
ウィキメディア・コモンズには、視覚に関連するカテゴリがあります。 |
外部リンク
- “ビジュアル生理学 - 視覚”. ビジュアル生理学. 2011年8月20日閲覧。
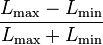 を用いることが多い。とは、画像中の輝度値の最大値と最小値を表す。この定義をMichelsonコントラストとよぶ。
を用いることが多い。とは、画像中の輝度値の最大値と最小値を表す。この定義をMichelsonコントラストとよぶ。




