ベーシック・ヘリックス・ループ・ヘリックス
- 関
- basic helix-loop-helix
Wikipedia preview
出典(authority):フリー百科事典『ウィキペディア(Wikipedia)』「2017/05/14 05:05:10」(JST)
[Wiki en表示]
| basic helix-loop-helix DNA-binding domain |
|
Basic helix-loop-helix structural motif of ARNT. Two α-helices (blue) are connected by a short loop (red).[1]
|
| Identifiers |
| Symbol |
bHLH |
| Pfam |
PF00010 |
| InterPro |
IPR001092 |
| SMART |
SM00353 |
| PROSITE |
PDOC00038 |
| SCOP |
1mdy |
| SUPERFAMILY |
1mdy |
| CDD |
cd00083 |
| Available protein structures: |
| Pfam |
structures |
| PDB |
RCSB PDB; PDBe; PDBj |
| PDBsum |
structure summary |
|
A basic helix-loop-helix (bHLH) is a protein structural motif that characterizes one of the largest families of dimerizing transcription factors.[2][3][4][5] It should not be confused with the helix-turn-helix domain.
Contents
- 1 Structure
- 2 Examples
- 3 Regulation
- 4 History
- 5 Human proteins with helix-loop-helix DNA-binding domain
- 6 References
- 7 External links
Structure
The motif is characterized by two α-helices connected by a loop. In general, transcription factors including this domain are dimeric, each with one helix containing basic amino acid residues that facilitate DNA binding.[6] In general, one helix is smaller, and, due to the flexibility of the loop, allows dimerization by folding and packing against another helix. The larger helix typically contains the DNA-binding regions. bHLH proteins typically bind to a consensus sequence called an E-box, CANNTG.[7] The canonical E-box is CACGTG (palindromic), however some bHLH transcription factors, notably those of the bHLH-PAS family, bind to related non-palindromic sequences, which are similar to the E-box. bHLH TFs may homodimerize or heterodimerize with other bHLH TFs and form a large variety of dimers, each one with specific functions.[8]
Examples
Examples of transcription factors containing a bHLH include:
- AhR
- Beta2/NeuroD1
- BMAL-1-CLOCK
- C-Myc, N-Myc
- MyoD
- Myf5
- Pho4
- HIF
- ICE1
- NPAS1, NPAS3, MOP5
- Scl, also known as Tal1
- proneural bHLH genes like p-CaMKII, and pSer(336)NeuroD.
- Scleraxis
- Neurogenins
- MAX
- OLIG1, OLIG2
- TCF4 (Transcription Factor 4)
- BHLHE41, also known as DEC2, SHARP1, and BHLHB3
bHLH transcription factors are often important in development or cell activity. BMAL1-Clock is a core transcription complex in the molecular circadian clock. Other genes, like c-Myc and HIF-1, have been linked to cancer due to their effects on cell growth and metabolism.
Regulation
Since many bHLH transcription factors are heterodimeric,[8] their activity is often highly regulated by the dimerization of the subunits. One subunit's expression or availability is often controlled, whereas the other subunit is constitutively expressed. Many of the known regulatory proteins, such as the Drosophila extramacrochaetae protein, have the helix-loop-helix structure but lack the basic region, making them unable to bind to DNA on their own. They are, however, able to form heterodimers with proteins that have the bHLH structure, and inactivate their abilities as transcription factors.[9]
History
- 1989: Murre et al. showed that dimers of various bHLH proteins bind to a short DNA motif (later called E-Box).[10] This E-box consists of the DNA sequence CANNTG, where N can be any nucleotide.[7]
- 1994: Harrison's[11] and Pabo's[12] groups crystallize bHLH proteins bound to E-boxes, demonstrating that the parallel 4-helix bundle motif loop orients the basic sequences to interact with specific nucleotides in the major groove of the E-box.
- 1994: Wharton et al. identified asymmetric E-boxes bound by a subset of bHLH proteins with PAS domains (bHLH-PAS proteins), including Single-minded (Sim) and the aromatic hydrocarbon receptor.[13]
- 1995: Semenza's group identifies hypoxia-inducible factor (HIF) as a bHLH-PAS heterodimer that binds a related asymmetric E-box.[14]
- 2009: Grove, De Masi et al., identified novel short DNA motifs, bound by a subset of bHLH proteins, which they defined as "E-box-like sequences". These are in the form of CAYRMK, where Y stands for C or T, R is A or G, M is A or C and K is G or T.[15]
Human proteins with helix-loop-helix DNA-binding domain
AHR; AHRR; ARNT; ARNT2; ARNTL; ARNTL2; ASCL1; ASCL2; ASCL3; ASCL4; ATOH1; ATOH7; ATOH8; BHLHB2; BHLHB3; BHLHB4; BHLHB5; BHLHB8; CLOCK; EPAS1; FERD3L; FIGLA; HAND1; HAND2; HES1; HES2; HES3; HES4; HES5; HES6; HES7; HEY1; HEY2; HIF1A; ID1; ID2; ID3; ID4; KIAA2018; LYL1; MASH1; MATH2; MAX; MESP1; MESP2; MIST1; MITF; MLX; MLXIP; MLXIPL; MNT; MSC; MSGN1; MXD1; MXD3; MXD4; MXI1; MYC; MYCL1; MYCL2; MYCN; MYF5; MYF6; MYOD1; MYOG; NCOA1; NCOA3; NEUROD1; NEUROD2; NEUROD4; NEUROD6; NEUROG1; NEUROG2; NEUROG3; NHLH1; NHLH2; NPAS1; NPAS2; NPAS3; OAF1; OLIG1; OLIG2; OLIG3; PTF1A; SCL; SCXB; SIM1; SIM2; SOHLH1; SOHLH2; SREBF1; SREBF2; TAL1; TAL2; TCF12; TCF15; TCF21; TCF3; TCF4; TCFL5; TFAP4; TFE3; TFEB; TFEC; TWIST1; TWIST2; USF1; USF2;
References
- ^ PDB: 1x0o; Card PB, Erbel PJ, Gardner KH (October 2005). "Structural basis of ARNT PAS-B dimerization: use of a common beta-sheet interface for hetero- and homodimerization". J. Mol. Biol. 353 (3): 664–77. doi:10.1016/j.jmb.2005.08.043. PMID 16181639.
- ^ Murre C, Bain G, van Dijk MA, Engel I, Furnari BA, Massari ME, Matthews JR, Quong MW, Rivera RR, Stuiver MH (June 1994). "Structure and function of helix-loop-helix proteins". Biochim. Biophys. Acta. 1218 (2): 129–35. doi:10.1016/0167-4781(94)90001-9. PMID 8018712.
- ^ Littlewood TD, Evan GI (1995). "Transcription factors 2: helix-loop-helix". Protein Profile. 2 (6): 621–702. PMID 7553065.
- ^ Massari ME, Murre C (January 2000). "Helix-loop-helix proteins: regulators of transcription in eucaryotic organisms". Mol. Cell. Biol. 20 (2): 429–40. doi:10.1128/MCB.20.2.429-440.2000. PMC 85097 . PMID 10611221.
- ^ Amoutzias, Grigoris D.; Robertson, David L.; Van de Peer, Yves; Oliver, Stephen G. (2008-05-01). "Choose your partners: dimerization in eukaryotic transcription factors". Trends in Biochemical Sciences. 33 (5): 220–229. doi:10.1016/j.tibs.2008.02.002. ISSN 0968-0004. PMID 18406148.
- ^ Lawrence Zipursky; Arnold Berk; Monty Krieger; Darnell, James E.; Lodish, Harvey F.; Kaiser, Chris; Matthew P Scott; Matsudaira, Paul T. McGill Lodish 5E Package - Molecular Cell Biology & McGill Activation Code. San Francisco: W. H. Freeman. ISBN 0-7167-8635-4.
- ^ a b Chaudhary J, Skinner MK (1999). "Basic helix-loop-helix proteins can act at the E-box within the serum response element of the c-fos promoter to influence hormone-induced promoter activation in Sertoli cells". Mol. Endocrinol. 13 (5): 774–86. doi:10.1210/mend.13.5.0271. PMID 10319327.
- ^ a b Amoutzias, Gregory D.; Robertson, David L.; Oliver, Stephen G.; Bornberg-Bauer, Erich (2004-03-01). "Convergent evolution of gene networks by single-gene duplications in higher eukaryotes". EMBO reports. 5 (3): 274–279. doi:10.1038/sj.embor.7400096. ISSN 1469-221X. PMC 1299007 . PMID 14968135.
- ^ Cabrera CV, Alonso MC, Huikeshoven H (1994). "Regulation of scute function by extramacrochaete in vitro and in vivo". Development. 120 (12): 3595–603. PMID 7821225.
- ^ Murre C, McCaw PS, Vaessin H, et al. (1989). "Interactions between heterologous helix-loop-helix proteins generate complexes that bind specifically to a common DNA sequence". Cell. 58 (3): 537–44. doi:10.1016/0092-8674(89)90434-0. PMID 2503252.
- ^ Ellenberger T, Fass D, Arnaud M, Harrison SC (April 1994). "Crystal structure of transcription factor E47: E-box recognition by a basic region helix-loop-helix dimer". Genes Dev. 8 (8): 970–80. doi:10.1101/gad.8.8.970. PMID 7926781.
- ^ Ma PC, Rould MA, Weintraub H, Pabo CO (May 1994). "Crystal structure of MyoD bHLH domain-DNA complex: perspectives on DNA recognition and implications for transcriptional activation". Cell. 77 (3): 451–9. doi:10.1016/0092-8674(94)90159-7. PMID 8181063.
- ^ Wharton KA, Franks RG, Kasai Y, Crews ST (December 1994). "Control of CNS midline transcription by asymmetric E-box-like elements: similarity to xenobiotic responsive regulation". Development. 120 (12): 3563–9. PMID 7821222.
- ^ Wang GL, Jiang BH, Rue EA, Semenza GL (June 1995). "Hypoxia-inducible factor 1 is a basic helix-loop-helix-PAS heterodimer regulated by cellular O2 tension". Proc. Natl. Acad. Sci. U.S.A. 92 (12): 5510–4. doi:10.1073/pnas.92.12.5510. PMC 41725 . PMID 7539918.
- ^ Grove C, De Masi F, et al. (2009). "A multiparameter network reveals extensive divergence between C. elegans bHLH transcription factors". Cell. 138 (2): 314–27. doi:10.1016/j.cell.2009.04.058. PMC 2774807 . PMID 19632181.
External links
- PDOC00038 in PROSITE
- Basic Helix-Loop-Helix Transcription Factors at the US National Library of Medicine Medical Subject Headings (MeSH)
- bHLH family at PlantTFDB:Plant Transcription Factor Database
UpToDate Contents
全文を閲覧するには購読必要です。 To read the full text you will need to subscribe.
English Journal
- Novel alternative splice variants of chicken NPAS3 are expressed in the developing central nervous system.
- Shin J, Kim J.SourceDepartment of Life Sciences and Ewha Research Center for Systems Biology, Ewha Womans University, Seoul 120-750, Korea.
- Gene.Gene.2013 Nov 10;530(2):222-8. doi: 10.1016/j.gene.2013.08.024. Epub 2013 Aug 18.
- We report isolation of novel splice variants of chicken Neuronal Per-Arnt-Sim domain protein 3 (cNPAS3) gene distinct from the previously predicted cNPAS3 at the 5' end. Newly identified cNPAS3 splice variants feature N-terminus coding sequences with high degrees of homology to human NPAS3 (hNAPS3).
- PMID 23962688
- A Novel Role of the STAT3 Pathway in Brain Inflammation-induced Human Neural Progenitor Cell Differentiation.
- Chen E, Xu D, Lan X, Jia B, Sun L, Zheng JC, Peng H.SourceUniversity of Nebraska Medical Center, 985930 Nebraska Medical Center, Omaha, NE 68198-5930, USA. jzheng@unmc.edu.
- Current molecular medicine.Curr Mol Med.2013 Nov;13(9):1474-84.
- Brain inflammation is a primary pathological driving force of many neurodegenerative disorders. In the destructive process, pro-inflammatory cytokines (IL-1β and TNF-α), are robustly released, affecting normal neural progenitor cell (NPC) differentiation, and resulting in a vast number of astrocyt
- PMID 23971732
- Increased lysosomal biogenesis in activated microglia and exacerbated neuronal damage after traumatic brain injury in progranulin-deficient mice.
- Tanaka Y, Matsuwaki T, Yamanouchi K, Nishihara M.SourceDepartment of Veterinary Physiology, Graduate School of Agricultural and Life Sciences, The University of Tokyo, Bunkyo-ku, Tokyo 113-8657, Japan.
- Neuroscience.Neuroscience.2013 Oct 10;250:8-19. doi: 10.1016/j.neuroscience.2013.06.049. Epub 2013 Jul 2.
- Progranulin (PGRN) is known to play a role in the pathogenesis of neurodegenerative diseases. Recently, it has been demonstrated that patients with the homozygous mutation in the GRN gene present with neuronal ceroid lipofuscinosis, and there is growing evidence that PGRN is related to lysosomal fun
- PMID 23830905
Japanese Journal
- Real-time imaging of bHLH transcription factors reveals their dynamic control in the multipotency and fate choice of neural stem cells
- Deubiquitinating enzymes regulate Hes1 stability and neuronal differentiation.
- CjbHLH1 homologs regulate sanguinarine biosynthesis in Eschscholzia californica cells.
Related Links
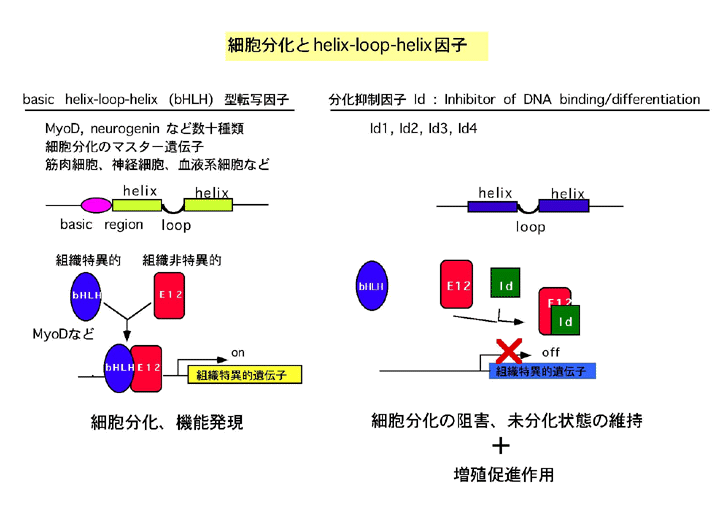
- 文献 [6] [7] による。 発現パターンによる分類 Class I 多くの組織に普遍的に存在 Tcf3 (E12/E47), Tcf4 (E2-2) etc. Class II 組織特異的に発現 MyoD, Neurog (Ngn), Hes etc. 組織特異的に発現するbHLH因子 (Class II ...
- はじめに bHLH(basic helix-loop-helix)及びPAS(Per-Ahr/Arnt-Sim)ドメインを持つ転写調節因子ファミリーはバクテリ アからヒトまで生物界で広く存在し,現在ヒトにおいて30 個以上単離されている。これらの転写調節因子ファミリー
Related Pictures






★リンクテーブル★
[★]
ベーシック・ヘリックス・ループ・ヘリックス
- 関
- bHLH
[★]
- 英
- basic helix-loop-helix、bHLH
[★]
- 同
- basic helix-loop-helix