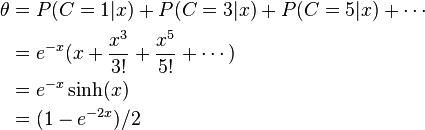遺伝的組換え(いでんてきくみかえ)は、狭義には、生物自身が遺伝子をコードするDNA鎖を途中で組み変える現象を差す。英語のRecombinationに相当する言葉として用いられる。広義には人工的な遺伝子組み換えも遺伝的組換えと記述される。
目次
- 1 組換え価
- 2 ホールデンのマップ関数
- 3 組換えホットスポット
- 4 疾患遺伝子マッピングにおける使用
- 5 DNAレベルの組換え
- 6 相同組換え
- 7 他のタイプの組換え
- 7.1 部位特異的組換え
- 7.2 非相同組換え
- 7.3 体細胞における組換え
- 8 脚注
- 9 関連項目
組換え価
同じ染色体上にある二組の遺伝子座間で組換えが起こる確率を組換え価(または組換え確率、英語: recombination rate)と言う。幾つかのモデル生物でゲノムの全塩基配列が決定された結果、遺伝的組換え価に基づいた遺伝学的地図上の距離と染色体上の物理的な距離とは必ずしも相関しないことが明らかになっているが、現在でも遺伝的組換えの結果を推定する際には有用な指標となっている。
片親(父親でも母親でもよい)由来の遺伝子座j上の対立遺伝子が、そのさらに父親(つまり祖父)由来であるとき、母親(つまり祖母)由来であるときであるとする。定義によれば、であるとき、遺伝子座jと遺伝子座j'の間に組換えが起こったと言う。従って組換え確率θは次のように定義される。
特定の家系サンプルにおいて、組換えが完全に観察できるとした場合、組換え価 (%) をl、組換えの起こった配偶子数をm、全ての配偶子数をnと置くと、組換え価は以下の式で推定できる。
組換え価l = 0%のときを完全連鎖、0% < l < 50%のときを不完全連鎖、l = 50%のときを独立という。染色体の乗り換えは、二本の染色体間のみで起こり、他の染色体間では起こらない。したがって、定義上組換え価が50%を超える事は有り得ない。
ホールデンのマップ関数
ここで組換え価をθとする。乗換えはまったくランダムに起きると考える。特定の二つの遺伝子座の間で乗換えが起こるというのはとても頻度の低い現象であると考えられるから、これがポアソン分布に従っているとする。この確率の期待値は上述の仮定によると、この遺伝子座の間の距離と相関していると考えられ、これをxとする。すなわち、c回の乗換えが起こる確率は以下のように表せる。
組換えが起こったということは、前述のように奇数回の乗換えが起こったことを示す。すなわち
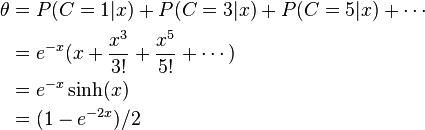
ここでsinhは双曲線関数の一つ。つまり
これはホールデンのマップ関数として知られ、二つの遺伝子座の間の遺伝的距離を表している。xの単位はモルガンMorgans (M)で、1Mの距離にある遺伝子座の間では1回の伝達につき1回の乗換えが生じることが期待される。これはきわめて長い距離に相当し、通常は1/100M=1cM(センチモルガン)の単位が頻用される。これを利用して染色体上の遺伝子の配置位置の遺伝子地図が書ける。
組換えホットスポット
ホールデンは上述のようにゲノム上のどこにおいても乗換えはランダムに起こると考えた。しかしこの仮定は誤っていたことがわかっているとすでに述べた。ゲノム上で、特に組換えが起こりやすい場所のことを組換えホットスポットと言う。
ヒトゲノムにおいては全ゲノム上における組換えホットスポット分布が確認されている。HapMap[1]の第二相データでは、ヒトゲノム上に32966個の組換えホットスポットが存在することがわかった[2]。特定の染色体により組換えホットスポットが多いということはない。
疾患遺伝子マッピングにおける使用
組換えは連鎖を弱めさせる主要な原因であり、組換えが存在するということそのものが連鎖解析を成立させている。組換え確率の正確な測定と疾患の伝達との関連を見ることにより、疾患の原因遺伝子は一意に特定されてもよいように思われる。しかし組換え確率の推定は前述のように不正確な要素も多く、また組換えホットスポットの存在によって結果として疾患関連領域の位置は大まかにしかわからない。現在現実の疾患遺伝子マッピングでは、むしろ連鎖不平衡のほうがより詳細な領域を同定するために有用である。
DNAレベルの組換え
分子生物学では、連続したDNA分子の中のある部分が切断と再結合により他のDNA分子の一部と混ぜ合わされることを組換えという。上記の染色体内での組換えはこのDNAの組換えによるものであるが、DNAの組換えには、他にもいろいろなタイプがある。
なお分子生物学あるいはバイオテクノロジーの基礎技術である人工的なDNA組換え(遺伝子組換えあるいは遺伝子工学)も組換えと呼ばれるが、仕組みは異なる。
相同組換え
染色体の組換えは普通、相同性のあるDNAの間で行われる。これを相同組換えという。
減数分裂の過程で染色体の乗換えに伴うのが普通であるが、体細胞分裂での乗換えに伴うものもある。
相同組換えであっても、染色体の別の位置(染色体レベルでは相同でない)の間で組換えが起これば、座位の数が変化する。その範囲に遺伝子が含まれていれば、遺伝子の重複または欠失につながる。これを不等組換えといい、不等乗換えに当たる。
これらの相同性があるDNA配列の間での組換え反応(相同組換え)を触媒するのは、組換え酵素(リコンビナーゼ)と呼ばれる酵素である。
大腸菌を含む真性細菌においてはRecAと呼ばれるリコンビナーゼが相同組換えを介してDNA修復や外来DNAの取り込みに関与している。一般にリコンビナーゼは細胞にとって重大な障害であるDNAの二本鎖切断(DSBs)の修復に重要である。大腸菌においては電離放射線やDNA複製の失敗によってDNAの二重鎖が切断されると、RecBCDと呼ばれるヘリカーゼとヌクレアーゼの複合体によりその末端の認識・消化が行われ一本鎖DNAが生じる。通常、生体内の一本鎖DNA領域は一本鎖結合蛋白質 (ssDNA binding protein, SSB) によって保護されているがRecBCDの働きにより一本鎖DNA上にRecA蛋白質が配置される。その後、RecAタンパク質がSSBを除去しながら一本鎖DNA上に重合・伸長することにより右巻き螺旋のヌクレオプロテインフィラメントが形成される。その後RecAフィラメントは染色体上の相同領域を探しあて組換え反応を行う。
酵母やヒトを含む真核生物では2種のリコンビナーゼが知られている。そのうちRad51タンパク質は体細胞分裂および減数分裂での相同組換えに必要である。もう一つのDmc1タンパク質は減数分裂時の相同組換えに特異的に機能する。真核生物ではDNA二重鎖の切断末端はMre11/Rad50/Nbs1 (Xrs2) 複合体によって認識され、さらにこの複合体を中心にその後の修復様式が制御されている。相同組換えを介した修復が行われる際は、ヘリカーゼおよびヌクレアーゼによって一本鎖領域が生じ、さらに一本鎖DNA結合蛋白質 (RPA) によって安定化される。その後Rad52タンパク質等の組換え触媒蛋白質が一本鎖DNA上でRPAを除去しRad51を配置することで、最終的にRecAと同様のヌクレオプロテインフィラメントを形成し相同組換え反応を起こす。
他のタイプの組換え
部位特異的組換え
バクテリオファージ(ファージ)による部位特異的組換えでは、ファージのDNAが乗換えと似た方法で、宿主である細菌の染色体DNAに組み込まれる。ファージDNAの部分配列が標的DNAのそれと完全に一致すると、インテグラーゼと呼ばれる特殊な組換え酵素がファージDNAを標的に組み込む。このようにファージDNAが染色体に組み込まれた状態をプロファージという。
非相同組換え
以上と異なり、相同性のないDNA配列の間での組換え(非相同組換え)もある。プロファージDNAが染色体から抜け出す(切り出されて新しいファージを作る)際にこれが起きることがあり、この場合には宿主の遺伝子を含んだファージ(形質導入ファージ)ができる。
体細胞における組換え
体細胞では、染色体の乗換えに伴う組換えとして、無性生殖する生物におけるものがある。 またDNA損傷が完全に修復されないまま細胞分裂を経た場合に起きる姉妹染色分体交換(Sister chromatid exchange:SCE)なども知られる。
そのほかに生理的に起きる組換えも知られており、代表的なものがリンパ球のB細胞における免疫グロブリン(抗体)遺伝子や、T細胞における受容体遺伝子である。これは抗体や免疫反応における機能的多様性に必須の過程である。
脚注
- ^ HapMap 計画
- ^ International HapMap Consortium. A second generation human haplotype map of over 3.1 million SNPs. Nature 2007;449:851.
関連項目